EFFECT OF Zn AND Mn SOURCES IN MICRODIETS ON GROWTH PERFORMANCE AND BONE DEVELOPMENT OF GILTHEAD SEABREAM
Introduction
Microdiets are being used to partially replace live feed in Gilthead seabream (Sparus aurata) larvae. However information on requirements of micronutrients for this species is scarce. Skeletal anomalies are highly present in seabream culture, and prevalence of severe anomalies ranges between 15-100% in commercial hatcheries. A wide range of factors are involved in the presence of skeletal anomalies including genetical, physical, chemical and nutritional. Zinc (Zn) and manganese (Mn) deficiencies have been related to reduced performance and higher prevalence of skeletal anomalies (Watanabe et al., 1997). However microdiets employed in larval nutrition mean the feed is more exposed to leaching processes. This, added to the fact that the larval digestive system isn't fully developed suggest that the source of Zn and Mn used may influence the bioavailability of the mineral.
Materials and Methods
Diets. Seven different isoenergetic and isonitrogenous practical diets were designed to contain Zn and Mn in either organic (Zn O and Mn O), inorganic (Zn I and Mn I) or encapsulated form (Zn E and Mn E) or none of these (Control -).
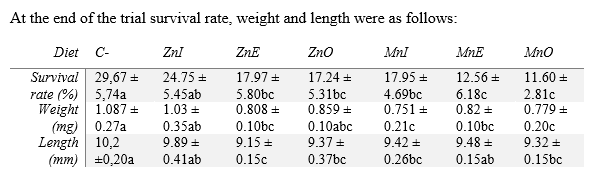
Fish and experimental conditions. Larvae from natural spawns from the broodstock of Aquaculture Research Group (GIA) (Las Palmas de Gran Canaria, Spain) with initial total length 6.4±0.3mm, dry body weight 0.22±0.01mg, mean ±SD previously fed rotifers (Brachinous plicatilis) enriched with DHA Protein Selco (INVE, Dendermond, Belgium) until 20dph, were randomly distributed in 18 experimental tanks at a density of 2100 larvae tank-1 and were manually fed one of the experimental diets tested in triplicates for 24 days, at a water temperature of 18.1-0.1ºC. All tanks (200 L light grey color cylinder fibreglass tanks) were supplied with filtered seawater (37 g/L salinity) at an increasing rate of 0.4-1 L/min along the feeding trials. Water entered the tank from the bottom and came out from the top; water quality was daily tested, and no deterioration was observed. Water was continuously aerated (125 mL/ min) attaining 5-8 g/L dissolved O2, and saturation ranged between 60 and 80%. At the end of the trial total survival rate was calculated. 30 larvae per tank were weight and measured and productive parameters recorded. 50 larvae per tank were sampled and stored at 10% formaldehyde for posterior osteological analyses.
Osteology. Larvae were double stained with Alcian blue and alizarin red according to Socorro, (2006 after Taylor and Van Dyke, 1985 and Dingerkus and Uhler, 1977) and skeletal anomalies were studied and classified according to Boglione et al., (2001). Frequencies of individuals with at least one anomaly, and frequencies of individuals with at least one serious anomaly (cranial anomalies, kyphosis, lordosis, vertebrae fusion or deformation) in each group were calculated.
Statistics. All data were tested for normality and homogeneity of variances with Levene′s test, not requiring any transformation and treated using one-way ANOVA. Means were compared by Duncan's test (P < 0.05) using a SPSS software (SPSS for Windows 11.5; SPSS Inc., Chicago, IL, USA). Bone anomalies present in fish from the different diets were subjected to correspondence analysis in order to relate the role of each diet on the frequency of anomalies (Hammer et al 2001.) using a PAST software (version 2.14, available at: www.nhm.uio.no/norlex/past/ download.html).
Results
Osteological analysis revealed only cartilage was stained. Every individual evaluated presented skeletal anomalies. Prevalence of severe anomalies almost reached 100% in every tank due to the presence of anomalies in the branchiostegal rays and lordosis. No significant differences were found between treatments for total prevalence of skeletal anomalies or severe anomalies.
Discussion and conclusion
Knowledge about marine fish species regarding mineral requirements is scarce (Hamre et al., 2013). On the other hand, information regarding mineral sources for seabream is almost inexistent (Antony Jesu Prabhu, 2015. PhD thesis). A reduction in survival rate, length and weight of fish fed either Zn or Mn with any source compared to those fed a control diet was found. This results are similar to others obtained in similar studies and suggest a toxic effect of either Zn or Mn (Ghrab, 2014. Master thesis), however the previous study also included selenium and iron, thus suggesting these two minerals weren't exerting a toxic effect. Amongst growth parameters fish fed inorganic Zn presented higher performance suggesting a better absorption.
Osteological analyses demonstrated a reduced skeletal development evidenced by the absence of mineralized bone. Seabream larvae as small as 5.7mm (Faustino and Power, 1998) show mineralized bone, however these, being at least 9.15mm long, didn't. The background high prevalence of skeletal anomalies camouflaged the effects of mineral supplementation, however the high prevalence of anomalies may be due to both a toxic effect of Zn and Mn in the diets containing either mineral, or to deficiencies in other minerals in the control - diet.
References
∙βBoglione, C., Gagliardi, F. and Scardi, M. (2001). Skeletal descriptors and quality assessment in larvae and post-larvae of wild-caught and hatchery-reared gilthead sea bream (Sparus aurata L. 1758). Aquaculture 192, 1-22.
∙βDingerkus, G. and Uhler, L.D. 1977. Enzyme clearing of alcian blue stained whole vertebrates for determination of cartilage. Stain Technology 32, 229-232.
∙βFaustino, M. and Power, D.M. (1998). Development of osteological structures in the sea bream: vertebral column and caudal fin complex. J. Fish Biol. 52, 11-22.
∙βHammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4: pp 9.
∙βHamre, K., Yúfera, M., Rønnestad, I., Boglione, C., Conceição, L. E. C. and Izquierdo, M. (2013). Fish larval nutrition and feed formulation: knowledge gaps and bottlenecks for advances in larval rearing. Rev. Aquac. 5, S26-S58.
∙βTaylor, W.R., and Van Dyke, G.V. 1985. Revised procedures for staining and clearing fishes and other vertebrates for bone and cartilage studies. Cybium 9, 107-119.
∙βWatanabe, T., Kiron, V. and Satoh, S. (1997). Trace minerals in fish nutrition. Aquaculture 151, 185-207.