Use of genome-wide information in the creation of base populations for aquaculture breeding programs
Introduction
The success of an aquaculture breeding program critically depends on the way in which the base population of breeders is constructed since all the genetic variability for the traits to be selected for is contained in those initial founders. Factors involved in the construction of the base population which are known to affect the efficiency of the selection program include the population size, the number of strains involved, the genetic relationships within and between them, the genetic level for the trait of interest of the founders and the way in which they are mated to create the initial stock (Holtsmark et al. 2008a, b). Traditionally base populations were founded from wild strains, but for some aquaculture species improved strains are also available. Thus, the genetic value of each strain is also a key factor to account for when designing base populations.
The increasing availability of genomic resources in aquaculture species allows us to improve the design of base populations. Particularly, genome-wide information could help us to refine the estimation of genetic relationships within- and between populations and to optimise the percentage or number of individuals to be sampled from each strain. Also, information on the individual or strain average phenotypic or estimated breeding values could be used for choosing founders that make the program to start from high levels of performance.
The objective of this study was to evaluate the efficiency of different strategies for creating base populations using molecular information to capture the highest initial genetic diversity and accounting or not for the phenotypic performance for the trait to be selected in the subsequent breeding program.
Material and methods
Diploid individuals were simulated having a genome comprising 20 chromosomes of 1 Morgan each. Each chromosome carried 50,000 neutral biallelic loci evenly spaced. Additionally, 5 to 5,000 biallelic markers per chromosome were simulated interspersed of the neutral loci. The latter were used to optimise the composition of the base population while the neutral loci were used to monitoring the effect on diversity of each strategy.
To obtain a realistic pattern of linkage disequilibrium a large population (N =1,000 with equal number of males and females) was simulated during 1,000 generations under random selection and mating, until the expected heterozygsity of the population reached an equilibrium value. A trait with heritability 0.4 and phenotypic mean and variance of 100 and 30, respectively, was simulated. The trait was controlled by 1,000 randomly chosen loci.
From the equilibrium population ten different strains were randomly sampled. These strains differed in size (from 10 to 40 individuals, half of each sex) in order to simulate different degrees of genetic drift. Then, they were allowed to diverge during 20 generations under three different scenarios: i) random selection; ii) stabilizing selection with different optima in order to mimic local adaptations in the nature; iii) artificial selection with different selection pressures in order to mimic the existence of already improved strains. At the end of this period subpopulations were expanded to have a large enough number of candidates.
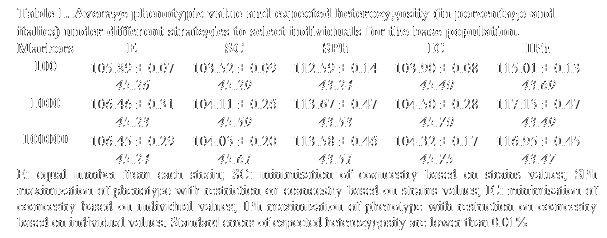
Base populations were constructed following several strategies: i) taking at random equal numbers of individuals from each strain (E); ii) determining optimal proportions to maximize the expected heterozygosity calculated from the mean coancestry values within and between strains (SC); iii) maximizing mean (strain) phenotypic value with a restriction on coancestry (SPh); iv) as in ii) but using individual relationships instead of strain averages (IC); v) as in iii) but maximizing individual values of the selected individuals instead of means (IPh). Comparisons between strategies were made in terms of the mean phenotypic value (M) and the expected heterozygosity (EH) of the group of selected individuals, as a measure of the genetic variation of the population. Values presented are the average of 30 replicates.
Results and Discussion
As can be seen in Table 1 the highest values of diversity are obtained with strategy IC and the highest means for the trait are obtained with strategy IPh. Under IPh, an increase in performance of 12% can be obtained with just a 5% reduction in the maximum diversity. Therefore, it seems that a compromise solution between diversity and performance can be found when creating base populations.
Values for both M and EH obtained when relying on strain averages are less than 2% lower than those observed when individual genotypes of candidates are available. This is an appealing result for cases where the budget is low and no all candidates can be genotyped. Moreover, scenarios with a limited number of markers (i.e. 100) imply only a loss of 1% from results with larger numbers.
Consequences of each strategy on the evolution of subsequent breeding programs, regarding short- and long-term responses and the accumulation of inbreeding need to be studied.
References
M. Holtsmark, G. Klemetsdal, A. K. Sonesson, J. A. Woolliams. 2008a. Establishing a base population for a breeding program in aquaculture, from multiple subpopulations, differentiated by genetic drift: I. Effects of the number of subpopulations, heritability and mating strategies using optimum contribution selection. Aquaculture 274: 232-240.
M. Holtsmark, G. Klemetsdal, A. K. Sonesson, J. A. Woolliams. 2008b. Establishing a base population for a breeding program in aquaculture, from multiple subpopulations, differentiated by genetic drift: II. Sensitivity to assumptions on the additive genetic relationships of base animals. Aquaculture 274: 241-246