SPATIAL AND TEMPORAL DYNAMICS OF DISEASE-INDUCED MORTALITY OF PACIFIC OYSTERS ALONG THE FRENCH COASTLINE REFLECT LIFE HISTORY, LOCATION AND ENVIRONMENTAL FACTORS
Introduction
Since 2008, oyster spats (Crassostrea gigas) have been recurrently decimated in France. These mortality events are associated with the infection of oysters with a newly described genotype of ostreid herpesvirus 1 (OsHV-1). In 2012, important mortalities of adult oysters were also reported and associated with bacteria of the Vibrio genus, in particular Vibrio aestuarianus (Barbosa Solomieu et al., 2015). Oysters are grown in estuaries and bays where they are naturally exposed to contrasted and fluctuating environmental parameters (e.g. temperature, salinity, phytoplankton concentration). These environmental parameters influence host-pathogen interactions by acting directly on the pathogen, on the host, or on both, and may modulate the severity of the disease.
The main objective of this study is to examine environmental risk factors of disease-induced mortality on oysters deployed in main French farming.
Materials and methods
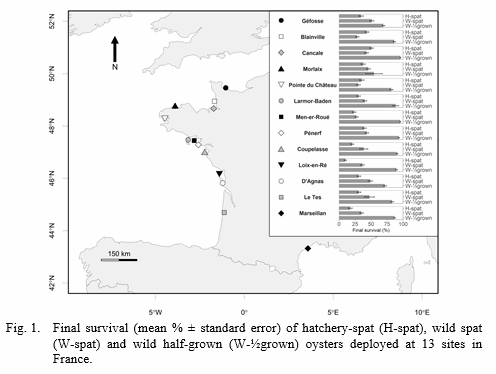
Three batches of sentinel oysters were deployed in February 2013 at 13 sites along French coasts (figure 1), and their survival was monitored bi-monthly from 11 March (t0) to 16 December (t280). The three batches of oysters consist of 3n spats originating from a private hatchery (H-spat), 2n wild spats (W-spat), and 2n wild half-grown oysters (W-½grown). The H-spat were initially free of OsHV-1 whereas the W-spat were asymptomatic carrier of the disease (Petton et al., 2015). Environmental (temperature, salinity, rainfall, chlorophyll a, concentration of phytoplankton cells, suspended particulate inorganic matter [SPIM], sea surface irradiance, wind speed) and host (DNA detection of OsHV-1 and V. aestuarianus) factors were recorded at the same time as survival. A logistic regression model was used to investigate the effect of sites and batches on the oyster final survival. The survival time curves of oysters were compared using a Cox regression model with sites and batches as fixed effect, and environmental factors (mean values and standard deviation measured before the onset of mortality) as covariates. A second Cox regression model was performed with level of OsHV-1 and V. aestuarianus DNA copy as time-dependent covariates.
Results
Final survival was 31.1 ± 11.1 % for H-spat, 39.7 ± 7.9 % for W-spat and 83.6 ± 11.9 % for W-½grown. Although sites and batches interacted in their effects on final survival, H-spats appeared the most susceptible (figure 1). Mortality initiated in W-spat followed by H-spat at every sites, reflecting that W-spat were asymptomatic carrier of the disease whereas H-spat were free of OsHV-1. Onset of mortality in H and W spat followed a latitudinal gradient from the South to the North. Suspended particulate inorganic matter (SPIM) and wind speed were negatively associated with the risk of mortality whereas chlorophyll a and OsHV-1 DNA were positively related with mortality risk.
Discussion and conclusion
Our study revealed that mortality of half-grown oyster was much lower than that of spat which were severely impacted by OsHV-1. Oysters that had survived a mortality event were thus naturally selected for resistance to OsHV-1 (Dégremont, 2011). Also, mortality initiated in W-spat followed by H-spat, reflecting the time-lag between disease expression in asymptomatic carrier of the disease and transmission of OsHV-1 particles to naïve individuals and expression of infection (Petton et al., 2015). The W-spat were generally more resistant than the H-spat, likely reflecting that they have been exposed to OsHV-1 during their first year on the field and partially selected for resistance. Finally, the onset of mortality in H and W spat followed a latitudinal gradient from the South to the North, which coincided with reaching the thermal threshold of 16°C in the seawater. Moreover, high level of chlorophyll a and low level of SPIM in seawater, two factors that reflect food availability, were associated with a higher risk of oyster mortality. In contrast, it was locally reported that well-fed oysters with high levels of energetic reserves are more resistant to OsHV-1 disease (Pernet et al., 2014). There is a need for more fundamental information on the nutritional physiology of the host in relation to viral infection. Overall, our results highlight the potential modulations of local-scale conditions on the kinetic and intensity of broad scale disease mortality events.
References
Barbosa Solomieu V., T. Renault, and M.A. Travers. 2015. Mass mortality in bivalves and the intricate case of the Pacific oyster, Crassostrea gigas. J. Invertebr. Pathol. 131: 2-10
Dégremont L. 2011. Evidence of herpesvirus (OsHV-1) resistance in juvenile Crassostrea gigas selected for high resistance to the summer mortality phenomenon. Aquaculture 317: 94-98
Pernet F., F. Lagarde, N. Jeanné, G. Daigle, J. Barret, P. Le Gall, C. Quere, and E.R. D'orbcastel. 2014. Spatial and temporal dynamics of mass mortalities in oysters is influenced by energetic reserves and food quality. PLoS One 9(2): e88469
Petton B., P. Boudry, M. Alunno-Bruscia, and F. Pernet. 2015. Factors influencing disease-induced mortality of Pacific oysters Crassostrea gigas. Aquac. Environ. Interact. 6: 205-222